Normal
0
false
false
false
false
EN-US
X-NONE
AR-SA
شناسایی و بررسی ساختار اجتماعات فولینگ مناطق زیستگاه مصنوعی سواحل بحرکان خوزستان
محقق : مجید شکاری(دکترای
زیست شناسی جانوران دریا و رئیس اداره شیلات شهرستان بهبهان)
چکیده: این مطالعه به منظور شناسایی و بررسی ساختار اجتماعات موجودات
چسبنده یا همان فولینگهای منطقه زیستگاههای مصنوعی با قدمت 13 سال، در سواحل
بحرکان واقع در شمالغربی خلیج فارس انجام شد. نمونهی فولینگ به صورت فصلی از بهار
1395 الی زمستان 395 از 4 ایستگاه به صورت
تصادفی با استفاده از کــوادرات cm 25×25 بوسیله
عملیات غواصی جمع آوری شدند. بر اســاس بیومس ( وزن تر ) گروههای مختلف چسبنده بر بدنه سازهها، از مرجانها رده Anthozoa با 88 درصد از بیومسکل، بیشترین توده
چسبنده را شامل شده و پس از آن اسفنجها (10 درصد) و
مرجانهای هیدروزوآ و بندپایان نیز هر یک با یک درصــد از مجموعه کـل را شامــل شدهاند. فصل بهار بیشترین میانگین فراوانی را نشان داده
است که ناشی از بیومس بالای گروه آنتوزوآ با 89 درصد در این فصل بوده است. بر اساس آزمون خوشهای دو فصل زمستان و پاییز
درصد تشابه بیشتری در حد 80 درصد داشته و
فصل تابستان و بهار درصد تشابه 50 درصدی را نشان می دهند.
کلمات کلیدی: زیستگاه مصنوعی،
موجودات چسبنده، بحرکان، مرجان، بیومس
مقدمه:
از زمانی که انسانها
شروع به استفاده از دریا و منابع آن کردهاند، ورود بسترهای سخت مصنوعی در دریا رو
به افزایش است که واضحترین و آشکارترین این فعالیتها، سازههای ساحلی است که به
طور چشمگیری طبیعت را تغییر میدهند (Glasby and Connell, 1999; Bacchiocchi and
Airoldi, 2003). همچنین تخریب و از بین
رفتن سواحل، زیستگاهها و اکوسیستمهای اقیانوسی یک مشکل و مساله جهانی می باشد، که
از زیستگاههای مصنوعی می توانیم برای بازسازی و ترمیم این اکوسیستمها استفاده
کنیم. برای قرنها نقش زیستگاههای مصنوعی در محیطهای آبی به تفضیل شرح داده
شده و کاربرد آن از مقیاسهای کوچک و استفاده از مواد طبیعی در ساخت آنها و افزایش
موفقیت در صید ماهی به صورت محلی، به سمت تکنولوژی پیچیده تر و استفاده از مدیریت
محیطی در مقیاس وسیع پیش می رود. کشورهای مختلف با توجه به مسائل و مشکلات خود
اهداف مختلفی را از ایجاد زیستگاههای مصنوعی دنبال می کنند. این ساختارها اغلب به
صورت مصنوعی و مطابق با پیش بینی تاثیر بر زندگی گونههای مورد نظر ساخته می شوند.
نقطه شروع کاربرد زیستگاههای مصنوعی از ژاپن بود که این ساختارها از منابع طبیعی
در مناطق گرمسیری و نیمه گرمسیری به کار برده شدند. اهداف تاریخی افزایش تولید غذا
هم در غالب ماهیگیری صنعتی (هند) و هم ماهیگیری تجاری (تایوان) ادامه یافت. از مهمترین
اهداف بیولوژیک این مطالعه، بررسی روند کلنی شدن ارگانیسمها روی سازههای مصنوعی می باشد. همچنین نوسانات
مکانی و زمانی تشکیل کلنی موجودات برای پیش بینی توسعه اجتماعات بنتیک وابسته به
تکیهگاه بر روی سازههای مصنوعی ضروری می باشد. مدلهای قدیمی تشکیل کلنی یک
توالی مستقیم را پیشنهاد می کنند که در آن گونههای پیشگام به سرعت و به تعداد
زیاد روی بستر تشکیل کلنی می دهند و به دنبال آن، اجتماعات با تنوع بالا به حالت
پایداری می رسند(Stanos
and Simon, 1980; Bailey-Brock, 1989). در حالی که Carter و Perkel در سال 2008 بیان کردند که مدل توالی قدیمی نمیتواند با
یک سیستم زیستگاه مصنوعی جدید ارزیابی شود. زیرا استقرار موجودات توسط ورود تصادفی
لاروها و غالبیت نسبی گونه های فرصت طلب تعیین می شود. علاوه بر این، پیچیدگی
بیولوژیکی زیستگاهها یک فاکتور مهم در ارتباط با نشست لاروها و شکلگیری اجتماعات
است. بنابراین تنها پس از گذشت یک زمان معین، گونههای دیگر هم شانس نشستن برروی
زیستگاه مناسب را پیدا می کنند در مطالعه اسکندری و همکاران (1387)، تعداد گروههای
جانوری چسبنده بیشتر بوده و شامل بارناکلها از سخت پوستان، مرجانها، بریوزوآ(
خزه شکلان)، خارپوستان، تونیکاتا، اسفنجها، پرتاران و نرمتنان بوده و بارناکلها
غالب بوده اند. بارناکلها که از موجودات پیشگام هستند دارای بیشترین فراوانی و
بیوماس در سالهای اول پس از احداث سازهها بودند، در حالی که درصد پوشش و تعداد
گونههای مرجان در منطقه خیلی کم بود(اسکندری و همکاران، 1387). با توجه به اهمیت
جوامع چسبنده در سری توالی جامعه جانوری در سازههای مصنوعی در این مطالعه سعی شد
به بررسی فرآوانی این گروه در زیستگاههای با قدمت 13 سال پرداخته شود تا با مقایسه با سازههای دارای عمر کمتر اهمیت
اینها بیشتر از پیش مشخص گردد.
مواد و روش ها:
زیستگاههای مصنوعی در منطقهی بحرکان با موقعیت جغرافیایی
17° 49 طول شرقی و 54 °29 عرض
شمالی در جنوب استان خوزستان واقع شده است. نمونه برداری به صورت فصلی از بهار 1395 الی زمستان
1395 صورت گرفت. به طور کلی 4 ایستگاه در نظر گرفته شد که شامل A,B,C,D هستند. مختصات ایستگاه ها در شکل 1 آمده است.
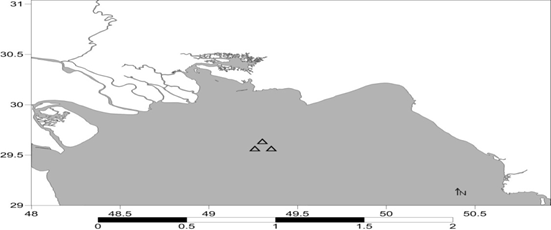
شکل1: موقعیت زیستگاههای
مصنوعی ایجاد شده در منطقه بحرکان خوزستان
به منظور بررسی اجتماعات کفزی نشست کرده روی بدنه سازهها،
نمونهبرداری با روش بررسی در محل استقرار توسط غوّاص و به صورت فصلی
صورت گرفت. در هر سازه از سه جهت راست، چپ و بالا نمونههایی در محدوده کوادرات (cm25×25) توسط غوّاص
با کمک کاردک و چکش خراشیده شده و به دورن کیسه نایلونی حاوی برچسب انتقال مییافت.
به عبارتی، در هر فصل از هر سازه سه نمونه به آزمایشگاه پژوهشکده آبزی پروری جنوب
کشور منتقل میشد. نمونهبرداری بهطور فصلی در طی یکسال از بهار 1395 لغایت زمستان1395صورت
گرفت. نمونههای موجود در بسته نایلونی پس از انتقال به بخش ساحلی به تفکیک
کوادرات محل مشاهده، ثبت شده و مورد جداسازی اولیه قرار گرفتند. سپس به ظروف مخصوص
برچسب زده شده حاوی الکل اتیلیک 90% انتقال داده شدند. نمونههای فیکس شده در جعبههای
بزرگ به آزمایشگاه انتقال یافت (Holme and Mcintyre, 1984).
در آزمایشگاه الکل اضافی نمونههای ماکروبنتوز را خارج
کرده و به بطریهای حاوی نمونه محلول رزبنگال 1 گرم در لیتر اضافه شد. پس از مدت 2
ساعت نمونهها از الک 5/0 میکرون عبور و رنگ اضافه شستشو داده شد. پس از شستشو،
نمونهها در بالاترین ردههای تاکسونومی ممکن جداسازی شدند و ماکروبنتوز موجود
مورد بررسی و جداسازی قرار گرفتند. نمونهها دربطریهای حاوی اتانول 90% قرار داده
شدند. سپس ازنمونههای توسط استریومیکروسکوپ متصل به دوربین عکسبرداری شد. با
استفاده از کلیدهای (Miner, 1950; Bayer et al.,
1983; Holthuis, 1985; Abele and Kim, 1986; Chace et al., 1986; Sterrer,
1986; Jones, 1986; Wing and; Colline et
al., 2005; Sanchez et al., 2005; Kenchington et al., 2009) تا سطح جنس یا در برخی موارد در سطح گونه
شناسایی شد. در این مرحله وزن تر نمونهها با ترازوی دیجیتال با دقت 001/0 گرم
برای ارائه بیومس تر توزین شد.
تراکم کفزیان بر اساس تعداد ارگانسیمها در هر متر مربع از
سطح مورد نمونهبرداری توسط فرمول 1 محاسبه گردید.
(فرمول 1) a× FD = i
که در آن i تعداد افراد جمعآوری شده
در هر کوادرات و a ضریب تبدیل به واحد مترمربع است.
دادههای بیوماس بر اساس وزن تر و واحد g/m2 بیان شد.
تجزیه و تحلیل آماری با استفاده از نرم افزار EXCEL و جهت تعیین انواع شاخصهای زیستی و آزمونهای چند متغیره ( تشابه، MDS، Anosim و آنالیز خوشهای ) از نرم
افزارهای Primer(5)
, و Biological toolsاستفاده شده است.
نتایج:
بر اساس بیومس( وزن تر)گروههای مختلف چسبنده
بر بدنه سازهها، از مرجانها رده Anthozoa با 88 درصد از بیومس کل، بیشترین توده چسبنده را شامل شده و پس از
آن اسفنجها (10 درصد) و مرجانهای هیدروزوآ و بندپایان نیز هر یک با یک
درصد از مجموعه کل را شامل شدند( شکل 2) شکل 3 میانگین فراوانی را در سازههای
مختلف را نمایش می دهد.
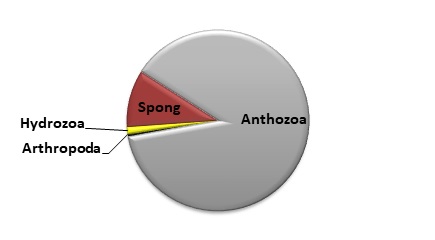
شکل 2: درصد گروههای
مختلف جانوران چسبنده به بدنه سازهها در سواحل بحرکان خوزستان(1395)
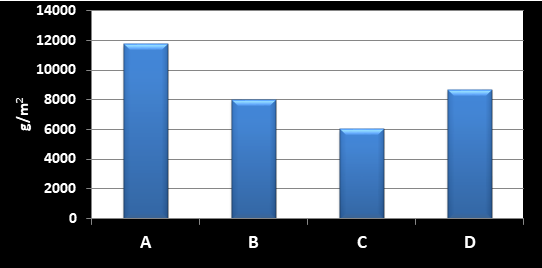
شکل 3: میانگین بیومس
جانوران چسبنده به بدنه سازهها در سواحل بحرکان خوزستان(1395)
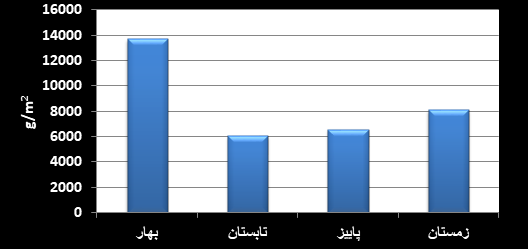
شکل4: میانگین بیومس
فصلی جانوران چسبنده به بدنه سازهها در سواحل بحرکان خوزستان(1395)
در شکل 3 میانگین بیومس
فصلی جانوران چسبنده بر بدنه سازهها در سواحل خوزستان نمایش داده شده است. فصل
بهار بیشترین میانگین فراوانی را نشان داده است که ناشی از بیومس بالای گروه
آنتوزوآ با 89 درصد در این فصل بوده است. در فصل تابستان نیز با وجودیکه درصد گروه
آنتوزوآ 89 درصد است اما بیومس کمتری را نشان داده است. در شکل 4 درصد بیومس گروههای
مختلف چسبنده به بدنه سازههای مختلف در فصول مختلف نمایش داده شده است.
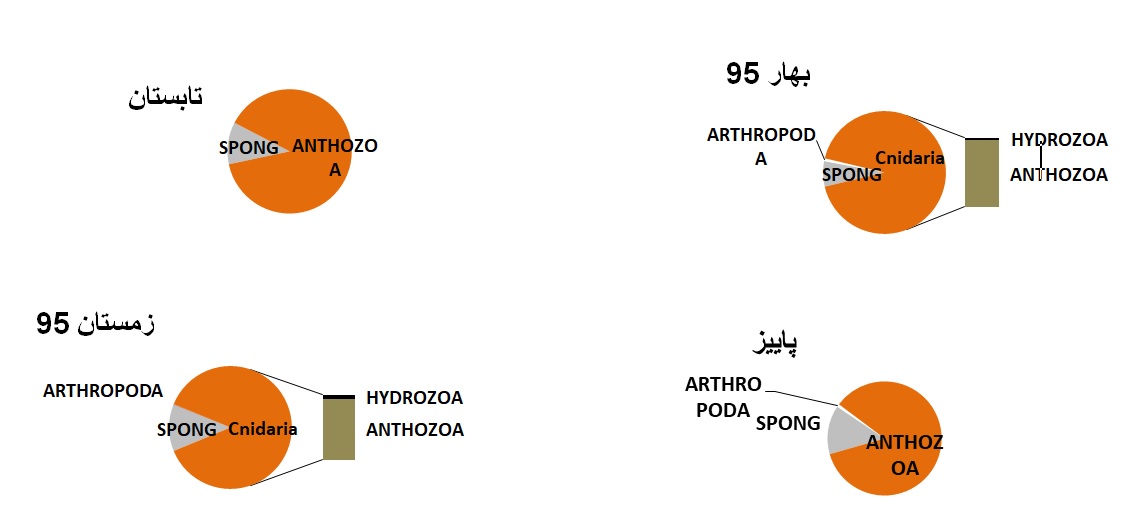
شکل5: نمایش درصد فراوانی حضورگروههای مختلف موجودات
چسبنده بر بدنه سازهها در فصول مختلف
جدول1: میانگین بیومس (
وزن تر/ گرم بر متر مربع) جانوران چسبنده بر بدنه سازهها در سواحل بحرکان خوزستان(1395)
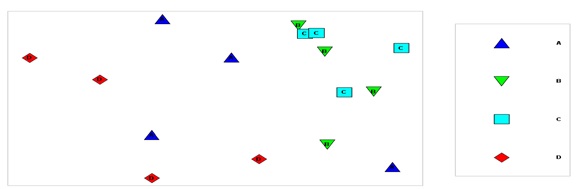
شکل6: آنالیز MDS بر اساس بیومس گونههای چسبنده در ایستگاههای مورد مطالعه (1395)
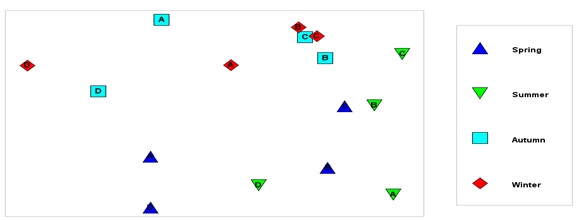
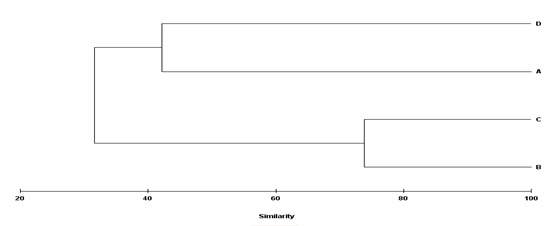
شکل7: آنالیز خوشهای بر اساس شاخص تشابه Bray-
Curtis بیومس گونههای چسبنده در فصول مختلف(1395)
بر اساس نتایج این آزمون
فصول پاییز و زمستان با تشابه بیشتر ی حدود 80 درصد بوده اند و دو فصل بهار و
تابستان درصد تشابه حدود 50 درصد را نشان داده اند. در شکل8- آزمون MDS بر اساس
بیومس گونه های چسبنده در سازههای مختلف ارائه شده است. تشابه Bray Curtis بر اساس (Log(x+1 داده ها
انجام شده است. فاکتور stress ، 0.12 محاسبه شده است. در شکل9 آنالیز خوشهای درصد شباهت نمونه های چسبنده در سازههای مختلف
را نمایش می دهد.
شکل8: آنالیز MDS بر اساس بیومس
گونه های چسبنده در ایستگاههای مورد مطالعه
شکل9: آنالیز خوشهای بر اساس شاخص تشابه Bray-
Curtis بیومس گونههای چسبنده در ایستگاههای مورد
مطالعه (1395)
بحث و نتیجه گیری
در مطالعه اخیر پس از
گذشت 13 سال از استقرار سازهها در دریا مشخص شد که مرجان ها از موجودات غالب بر
روی سازهها هستند. موجودات چسبنده از 3 گروه جانوری مرجانها(آنتوزوآ و هیدروزوآ)،
اسفنجها و سخت پوستان بوده اند و مرجانهای آنتوزوآ(Anthozoa) 88 درصد این مجموعه را شامل می شوند. علاوه بر
گونه Plumarella sp. ، سه گونه Ellisella barbadensis, Plexaura sp. و Eunicea sp. از
رده گورگونیاها بر روی سازهها مشاهده شد. گورگونیاها یک گروه مهم از مرجانهای
نرم در زیستگاههای آبسنگهای مرجانی در تمام دنیا هستند و مشخصه آنها کلنیهایی
با ظاهر شلاق مانند و باریک می باشد که از قاعده به بستر چسبیدهاند؛ و غالباُ
دارای انشعابات زیادی نیز هستند که در ستون آب گسترش می یابند. این مورفولوژی ویژه
باعث می شود که سطح وسیعی از کلنی در معرض جریان آب قرار گیرد. همچنین دامنه پراکنش و فراوانی این مرجانها
بستگی زیادی به فاکتورهای محیطی از جمله: نوع بستر، نور، دما، رژیم جریانات و سرعت
جریانات دارد (Kinzie, 1979; Weinberg, 1979). از میان فاکتورهای گفته شده بستر مهم ترین فاکتور محدود کننده
برای گورگونیاها و مرجانهای شاخهای می باشد؛ زیرا آنها در بستر های سخت جایی که
پوشش جلبکی حداقل است رشد می کنند. پس می توان علت حضور بالای این مرجانها در
منطقه را به خاطر حضور این بستر جدید در منطقه دانست. همچنین به دلیل ثبت مشاهدات
عینی غواص مبنی بر کدورت بالای محیط می توان نتیجه گرفت به دلیل اینکه گورگونیاها
و Sea whip ها مقاوم ترین گونه ها نسبت به کدورت و
رسوبات هستند، در منطقه به صورت غالب درآمده اند.دیگر محققان در بررسی های انجام
شده بر روی بسترهای مصنوعی با چنین شرایطی، مرجان های شاخهای (گورگونیا) را به
عنوان مرجان غالب معرفی کردند(Perkol-Finkel
and Benayahu, 2009; Clark and Edwards, 1999; Zeevi Ben-Yosef and Benayahu,
1999)
Lasker (
1998 ) بیان کرد که Octocoral ها دارای مکانیسم های تکثیرغیر جنسی سریع و
فعالی هستند که علت افزایش سریع جمعیت شان می باشد. همچنین دارای دوره لاروی
کوتاهی هستند که سریعا پس از آزاد شدن از بدن والدین ساکن می شوند. پس میتوان
نتیجه گرفت که این ویژگی بیولوژیکی ویژه باعث رشد انفجاری Octocoral ها و به خصوص گونه Plumarella sp. شده است.
همچنین میتوان پیش بینی کرد که در آینده نیز ممکن است تراکم بیشتری داشته باشند.
میتوان گفت بیش ترین میزان بیوماس مرجانها در فصل تابستان ناشی از همزمانی با
فصل زادآوری(تولید مثل جنسی) میباشد (Zeevi Ben-Yosef and Benayahu, 1999). اسفنجها
با 10 درصد پوشش دومین گروه غالب چسبنده در این مطالعه بوده اند که در تمامی فصول
درصد حضور اسفنجها نسبتا یکسان بوده است. در مطالعه اسکندری و همکاران (1387)
درصد فراوانی اسفنجها بیشتر از مطالعه اخیر بوده است که احتمالا این کاهش درصد میتواند
در ارتباط با سن سازهها باشد که با غالبیت مرجانها، درصد حضور اسفنجها کاهش
یافته است. فراهم آوردن زیستگاههای کوچک و بزرگ برای محافظت از آنها در برابر
سایر شکارچیان (استتار)، فراهم آوردن غذا (به صورت مستقیم یا غیرمستقیم) برای گروههای
جانوری، در نهایت منجر به افزایش تولید ثانویه در چرخهی حیات خواهد شد. مثالهای
زیادی در ارتباط با همکاری اسفنجها با سایر گروههای جانوری وجود دارد. اغلب آنها
در ارتباط با نوع مورفولوژی (داخلی و یا خارجی) یا اسکلت اسفنجها میباشد. بعضی
نرمتنان رابطهی همغذایی با اسفنجها دارد.
یک درصد مجموعه چسبنده را
بارناکلها شامل می شوند که در مقایسه با سالهای گذشته در سیر توالی جوامع
چسبنده سازهها، بارناکلها جزو گروههای
اولیه هستند که بمرور زمان جایگزین شدهاند.
بارناکلها برای نشستن نیاز به تکیهگاه سخت دارند و در سالهای بعد بدلیل رشد و
کلنی شدن تودههای حجیم مرجانها و اسفنجها، دسترسی به بدنه سخت سازهها امکان
پذیر نبوده است.
مقایسه بیومس وزن تر بین
سازههای مختلف اختلاف معنیداری نشان نمیدهد(p>0.05) در حالیکه آنالیز خوشهای بر اساس بیومس
چسبندهها در سازههای مختلف نشان داده است که دو گروه مشخص شامل سازههای(A و(D و
( Bو (C در سطح تشابه 45 درصد جدا شده و دو سازه B و C حدود 80
درصد تشابه را نشان میدهند که بدلیل دو گونه Plumarella sp. و Haliclona simulans بوده که
در دو سازه B و C بیومس بسیار بالایی را نسبت به سازههای
دیگر داشتهاند. در مطالعه اسماعیلی و همکاران (1391)، در سال مطالعه 1388، بطور مشخص سازه قدیمی D) ) با سایر سازهها، اختلافات مشخصی را در آنالیزهایMDS و
آنالیزهای خوشهای بر اساس میزان تشابه ترکیب گونهای چسبندهها در سازههای مختلف
نشان داده است که علت آن حضور بالای ماکروجلبکها در این سازه بوده است. ماکروجلبکها
در مطالعه اخیر حضور نداشتهاند.
در مطالعه اخیر فصلها
نیز اختلاف معنی داری در بیومس نشان ندادهاند. در حالیکه فصل بهار بیومس بیشتری نسبت به سایر فصلها
داشته است و آنالیز خوشهای بر اساس تشابه ترکیب گونهای فصول مختلف، فصول پاییز و
زمستان با بیشترین نسبت تشابه از دو فصل بهار و تابستان مجزا شده است. در مطالعه
اسماعیلی و همکاران (1391) نیز فصل بهار و تابستان، مشخصا با فصول پاییز و زمستان
بر اساس تشابه ترکیب گونهای جانوران چسبنده جدا شدهاند. این مورد احتمالا با دوره مناسب تولیدمثلی گروههای
چسبنده در بهار و تابستان و همچنین با رابطه تغذیه ای و فشار شکار ماهیان و آبزیان
منطقه مرتبط است.Smiley (
2006 )
بیان داشت که تغببرات فصلی شرایط محیطی بر تعداد و پراکنش گونههای مختلف و
اجتماعات موجودات چسبنده روی سازهها اثرگذار است. بر اساس یک تئوری اکولوژیکی در
محیطهای گرمسیری گونهها و نوسانات آنها کمتر به فصول وابسته است. آنچه در این
مناطق سبب غالب شدن برخی گونهها در فصول مختلف میشود، اندرکنش های بیولوژیکی
است. در حالی که در محیط های معتدله نوسانات فصلی در فاکتورهای فیزیکی در غالب
شدن اجتماعات اثرگذارند(Menge
and Sutherland, 1987).در جنوب شرق فلوریدا نیز این صخرههای مصنوعی بکار
گرفته شدند. مطالعات بلند مدت بر روی آنها نشان داد که 9 ماه پس از استقرار این
صخرهها اولین گروهی که بر روی صخرهها جایگزین شدند جلبکها بودند (Turf algae)، پس از آن بریوزوئنها، تونیکاتا، و هیدروئیدها در
منطقه جانشین شدند. البته جلبکها در تمام مدت مطالعات گروه غالب بودند. پس از پایداری
اکوسیستم مرجانهای آهکی نیز در منطقه استقرار یافتند و تنوع گونهها در منطقه به
طور چشمگیری نسبت به قبل از استقرار صخرهها و در چند ماهه اول استقرار افزایش
یافت که این افزایش تنوع با ثبات و پایداری اکوسیستم نیز بیشتر شد(Santos et al., 2011).پس از گذشت حدود 13 سال
از احداث سازهها در سواحل خوزستان، توالی گروههای مختلف در سازهها مشاهده شده و
بنظر به مرحله ثبات نزدیک میشود(اسکندری و همکاران، 1387و اسماعیلی و همکاران ،
1391) در مقایسه با موجودات اینفونا قبل از احداث سازهها، اپیفونا در سازههای
مصنوعی در خلیج Delaware از 147 تا 895 برابر
بیشتر بوده است(Foster
et al., 1994). در مطالعه
اخیربه دلیل کاهش فضای اشغال نشده و با توجه به
تغییرات کمی که در ترکیب گونهای مرجانها در فصلهای مختلف مشاهده شد میتوان
نتیجه گرفت که با گذشت زمان و افزایش عمر سازه از شدت نابودی و کلنی شدن اولیه
کاسته میشود و اجتماعات زیستی روی سازهها از ثبات بیشتری برخوردار خواهند شد.
منابع
اسکندری، غ.، دهقان مدیسه، س.، اسمائیلی، ف.،
سبزعلی زاده، س.، خلفه نیلساز، م.، صفی خانی، ح.، کاشی، م. میاحی، ی.، اژدری، ح. و
حسینی، س.، 1387. بررسی ساختار جمعیتی زیستگاههای مصنوعی احداث شده در سواحل
خوزستان. سازمان تحقیقات و آموزش کشاورزی. گزارش نهایی پروژه تحقیقاتی. 139 ص
اسماعیلی، ف.، دهقان مدیسه، س.، سبزعلی زاده،
س.، اسکندری، غ.ر.، کیان ارثی، ف.، میاحی، ی. و بنی طرفی،ج.1391. پایش زیستگاههای
مصنوعی احداث شده در سواحل خوزستان .
مؤسسه تحقیقات و آموزش شیلات ایران.94ص.
Abele, L. G.,
and Kim, W., 1986. An illustrated guide to the marine decapod crustaceans of
Florida. State of Florida department of environmental regulation. Vol: 8(1) part
1 and 2.
Bacchiocchi,
F. and Airoldi, L., 2003. Distribution and dynamics of epibiota on hard
structures for coastal protection. Estuarine, Coastal and Shelf Science, 56:
1157-1166.
Bailey-Brock, J.H., 1989. Fouling community
development on an artificial reef in Hawaiian waters. Bulletion of Marine
Science, 44: 580-591
Bayer, F.M.,
Grasshoff, M. and Verseveldt, J., 1983. Illustrated trilingual glossary of
morphological and anatomical terms applied to Octocorallia. 75p.
Carter, A. and
Prekel, S., 2008. Benthic colonization and ecological successional patterns on
a planned nearshore artificial reef system in Broward Country, SE Florida. In
Proceeding of the 11th International Coral Reef Symposium, Florida, pp.
1209-1213.
Clark, S. and
Edwards, A.J., 1999. An evaluation of artificial reef structures as tools for
marine habitat rehabilitation in the Maldives. Aquatic Conservation: Marine and
Freshwater Ecosystems, 9: 5–21.
Chace Jr., F.
A., McDermott, J. J., McLaughlin and Manning, R. B., 1986. Decopoda. In:
Sterrer, W. G., and Schoepfer- Sterrer, C., Marine fauna and flora of Bermuda a
systematic guide to the identification of